通过细菌和肿瘤细胞基因工程实现级联增强抗肿瘤免疫的近红外-II-反应性混合系统
摘要
在癌症免疫治疗的创新探索中,将纳米技术与肿瘤靶向细菌相结合的策略展现出了突破传统治疗局限性的巨大潜力。这一前沿方法旨在解决纳米粒子在生物体内蓄积不足、穿透深度有限及分布不均等关键问题,通过构建一种智能的混合治疗平台,实现了对免疫级联反应的显著增强,进而推动癌症免疫疗法的进步。
具体而言,本研究精心设计了一种集成了pH响应性席夫碱键连接机制与第二近红外(NIR-II)光热纳米粒子的工程化细菌混合系统。该系统巧妙地利用了细菌对肿瘤缺氧微环境的特异性靶向能力及其出色的组织穿透力,确保混合体能够高效、精准地聚集于肿瘤病灶区域。一旦到达目标位置,通过特定的pH环境变化,光热纳米粒子将从细菌表面解离,独立执行其多重治疗功能。
纳米粒子的独立行动首先聚焦于肿瘤细胞的基因工程改造,通过精准调控,诱导肿瘤细胞发生凋亡,并有效下调程序性细胞死亡配体1(PD-L1)的表达水平。这一过程不仅直接削弱了肿瘤细胞的生存能力,还通过降低免疫抑制性肿瘤微环境的屏障,为后续的免疫应答铺平了道路。
与此同时,NIR-II光热纳米粒子在温和的光照条件下产生的局部热效应,不仅促进了肿瘤相关抗原的释放,增强了免疫系统的识别与攻击能力,还触发了细胞因子如白细胞介素-2(IL-2)的持续表达。IL-2作为关键的免疫调节因子,能够进一步激活并增强免疫细胞的抗肿瘤活性,形成正向的免疫级联反应。
尤为值得关注的是,本研究还实现了p53基因转染与NIR-II光激活细菌基因工程之间的协同作用,这一创新点极大地提升了抗肿瘤效果,展现了混合系统在癌症治疗中的综合优势。综上所述,本研究不仅提出了一种构建高效混合治疗系统的创新策略,更为级联增强癌症免疫疗法的发展开辟了新的途径,预示着未来癌症治疗领域的重大突破。
1 引言
在癌症治疗领域,具备优异理化特性的多功能纳米粒子展现出了巨大的潜力,然而,其在肿瘤组织中的低蓄积性、有限的穿透力及分布不均等问题,显著制约了其抗肿瘤效果。与此同时,新兴的肿瘤靶向细菌以其独特的主动迁移能力,能够精准定位至缺氧且免疫抑制的肿瘤区域,实现深度渗透与均匀分布,为癌症治疗开辟了新径。这些细菌不仅具备特异性靶向特性,还展现出自我推进及在肿瘤环境中稳定增殖的非凡能力,加之灵活的生物工程技术与简便的化学修饰手段,使得它们成为抗肿瘤治疗的理想载体。
近年来,通过细菌高效递送纳米粒子至肿瘤部位,在癌症强化治疗方面取得了显著突破,这一策略巧妙结合了活微生物与纳米技术的互补优势,旨在减少治疗副作用的同时,显著提升治疗效果。其中,光热纳米粒子作为非侵入性光热疗法的关键元素,被广泛用于诱导肿瘤细胞死亡,并触发肿瘤相关抗原(TAAs)的释放,进而激活机体免疫反应。尤其是当这些纳米粒子搭载于肿瘤靶向细菌上时,能够在肿瘤区域富集,实现光热免疫疗法的有效实施。
然而,传统近红外光(如808纳米波长)在组织中的穿透力有限,且高温环境可能对免疫细胞造成损害,从而限制了治疗效果。相比之下,第二近红外(NIR-II)波段的光诱导光热效应更为理想,它能够实现更深层次的穿透,同时保持较低的副作用水平。特别是温和加热(约43℃)被认为能有效提升免疫细胞活性,诱导免疫原性细胞死亡(ICD),促进TAAs的释放,进而优化免疫治疗效果。
基于上述发现,将缺氧靶向细菌与光热纳米粒子整合为一个混合系统,利用NIR-II光诱导温和加热,以增强癌症免疫疗法,成为了一个极具吸引力的策略。此外,通过基因工程技术,细菌可持续表达治疗性蛋白或免疫调节剂,如白细胞介素-2(IL-2),后者已被FDA批准用于促进细胞毒性T淋巴细胞(CTL)和自然杀伤细胞(NK)的活化与增殖。控制免疫调节剂在肿瘤部位的精准表达,对于提高治疗效果、降低全身毒性至关重要,而这一过程可通过NIR触发的光热纳米粒子进行远程调控。
进一步地,肿瘤细胞的原位基因工程也为癌症治疗提供了新视角。例如,通过基因载体将p53抗原导入肿瘤细胞,可诱导其凋亡,并下调PD-L1的表达,从而增强免疫细胞对肿瘤细胞的识别与攻击能力。将细菌与光热纳米粒子相结合的混合系统,不仅能够持续表达免疫调节剂,还能递送p53基因,通过诱导ICD、表达免疫调节剂及下调PD-L1等多重机制,协同增强免疫级联反应。
尽管前景广阔,但构建这样一个复杂的混合系统以实现NIR-II光触发的细菌基因工程、ICD诱导及肿瘤细胞基因工程,仍面临诸多挑战。为此,作者提出了一种创新的混合治疗平台设计方案,该平台由工程菌与拉普兰/金纳米颗粒组成,通过基因工程技术对细菌和肿瘤细胞进行双重改造,旨在实现免疫级联增强的癌症治疗。该平台的核心在于近红外-II光响应的光热拉普兰/金纳米粒子,其独特的结构设计使得在NIR-II光照射下能够产生温和加热,进而触发一系列免疫激活与肿瘤细胞凋亡机制。通过体外与体内实验验证,作者证明了该混合平台在提高癌症免疫疗法疗效方面的可行性与有效性。
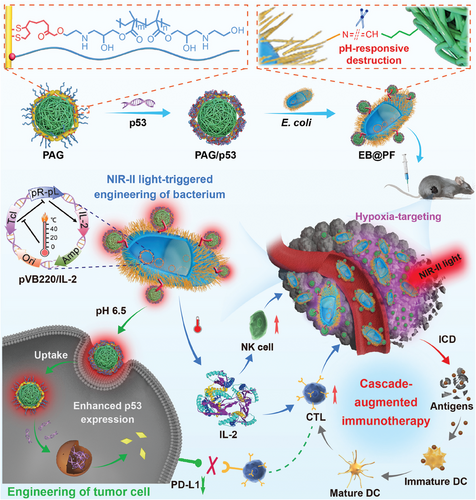
图 1 EB@PF 混合物的构建及其在通过缺氧靶向和近红外 II 光响应细菌和肿瘤细胞基因工程增强癌症免疫疗法中的应用示意图。
2 结果与讨论
2.1 细菌-纳米颗粒杂交平台的制备
在探索癌症治疗新策略的过程中,作者成功构建了一种集细菌与纳米技术于一体的创新杂交系统。该系统通过在非侵袭性大肠杆菌MG1655表面共轭光热纳米颗粒,实现了对细菌的光热响应性基因工程编程。具体而言,作者采用了包含热敏感启动子(pBV220)和免疫调节剂IL-2目的基因(pR-pL)的质粒,确保在温度高于42°C时,IL-2能够特异性表达。通过酶联免疫吸附试验(ELISA)验证,该系统在温度升高时显著提升了IL-2的表达水平,成功制备了热敏工程菌(EB)。
为了进一步提升治疗效果,作者制备了具有特定纵向表面等离子体共振(LSPR)峰的Au NRs,其波长约为1050 nm,并通过透射电子显微镜(TEM)确认了其形貌特征。随后,利用拉普兰纳米球与Au NRs的巧妙结合,合成了Pul-Au纳米粒子,并通过能量色散光谱(EDS)等手段验证了其结构组成。为了增强纳米粒子的基因递送能力,作者引入了阳离子聚合物PGEA,通过Au-S键将其共轭到金纳米棒表面,生成了Pul-Au-PGEA(PAG)纳米颗粒。这种新型基因载体不仅具有低细胞毒性和高转染性能,还显著提升了纳米粒子的生物相容性。
为了验证PAG纳米颗粒的基因递送能力,作者使用了编码雷尼拉荧光素酶的报告质粒pRL-CMV进行研究。结果显示,PAG/pRL-CMV复合物(PAP)具有适中的尺寸和正表面电荷,有利于细胞吸收和基因转染。随后,作者将PAP复合物与EB混合,通过静电相互作用和pH响应性席夫碱键构建了EB@P杂交体。这种杂交体在酸性肿瘤微环境(TME)中能够响应性释放PAP纳米颗粒,为肿瘤细胞的基因工程提供了可能。
综上所述,作者设计的这种细菌和纳米颗粒杂交系统,通过光热响应性基因编程和基因递送能力的双重提升,为癌症治疗提供了一种新颖而有效的策略。该系统不仅能够在肿瘤部位实现精准治疗,还能够通过调节免疫环境来增强治疗效果,展现出广阔的临床应用前景。
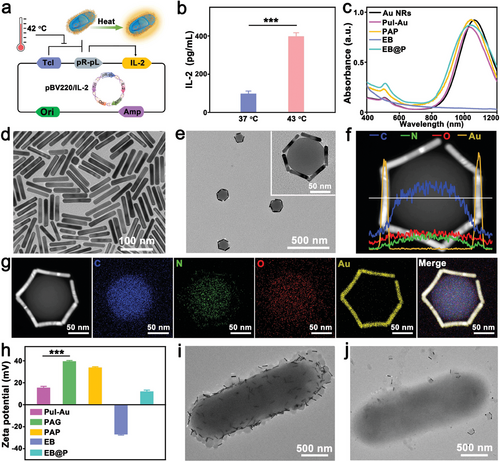
图 2 a) 基于质粒 pBV220 的 IL-2 表达机制。 b) 通过 ELISA 法检测 EB 的 IL-2 定量表达(平均值 ± SD,n = 3)。d) Au NRs 和 e) Pul-Au 纳米粒子的 TEM 图像。 f) Pul-Au 纳米粒子的 STEM 图像和 EDS 图谱。h) Pul-Au、PAG、PAP、EB 和 EB@P 的 Zeta 电位值(平均值 ± SD,n = 3)。**P<0.001,采用Student's检验。
2.2 细菌和肿瘤细胞的近红外-II 光响应基因工程
在深入探索EB@P(工程细菌与纳米颗粒的杂交体)的光热效应及其在治疗应用中的潜力时,作者系统评估了其在近红外-II波段(特别是1064纳米波长)的光照下的性能表现。实验结果显示,EB@P悬浮液的温度上升显著,这一变化与浓度、功率密度及照射时间紧密相关(图3a及图S5a),证明了其在光热治疗中的高效性。特别是在0.6 W/cm²功率密度下,10分钟的照射足以使EB@P溶液温度接近43°C,这一温度范围既满足了温和光热加热的需求,又有效避免了对周围健康组织的潜在损害(参考文献53, 54)。此外,EB@P在经历多次光热循环后依然保持稳定的光热性能(图S5b),彰显了其作为高效光热制剂的巨大潜力。
关于EB@P的生物学活性,作者发现近红外-II光照射并未对细菌存活率及生长率造成显著影响(图3b,c),同时ELISA检测结果显示,光热刺激有效触发了IL-2的表达,且表达水平与热诱导相当(图S7),表明PAP纳米颗粒对IL-2的表达无显著干扰。
进一步,作者通过MTT试验评估了EB@P的生物安全性,结果显示在特定浓度范围内,其对HEK293细胞的毒性可忽略不计(图3d),验证了其良好的生物相容性。在基因传递性能方面,EB@P在酸性条件下(模拟肿瘤微环境)展现出增强的转染效率(图3e),且温和光热加热进一步提升了这一效率,这与先前研究中光热加热促进基因治疗的机制相吻合(参考文献55-58)。
为了直观展示EB@P的肿瘤基因治疗效果,作者采用EGFP质粒及p53功能质粒进行了实验。结果显示,在pH 6.5条件下,经温和光热加热的OEB@PAG表现出最高的EGFP表达效率(图3f),而Western印迹和免疫荧光检测则证实了p53基因在B16F10细胞中的成功转染及PD-L1表达的显著下调(图3g及图S10),这对于缓解免疫抑制性肿瘤微环境具有重要意义(参考文献45)。
在体外抗癌功效评估中,EB@PF杂交体在含血清培养基中展现出优异的稳定性(图S11, S12),且其光热转换效率高达31.3%(图S13c,d),与同类材料相当。细胞存活率实验表明,EB@PF在光热协同作用下显著降低了B16F10细胞的活力(图3h),而CRT转位实验则进一步证实了混合系统诱导免疫原性细胞死亡(ICD)的能力(图3i,j及图S14-S16),为肿瘤治疗提供了新的策略。
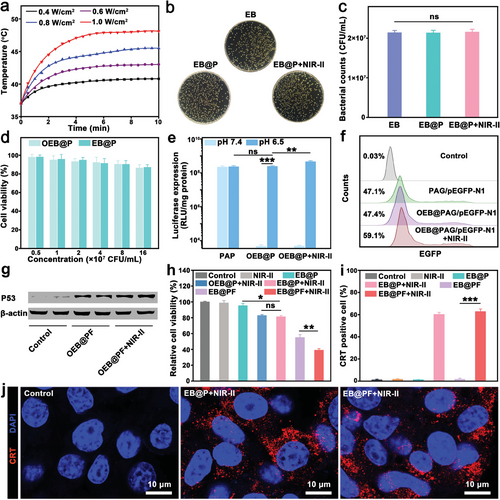
图 3
a) 不同激光功率密度下 EB@P 在培养基(2 × 10 7 CFU/mL)中的温升。 b) 用 NIR-II 激光(1064 nm, 0.d) 分别与不同浓度的 OEB@P 和 EB@P 培养的 HEK293 细胞的细胞活力(平均值±标 准差,n = 3)。 e) PAP、OEB@P 和 OEB@P 在 NIR-II 光照射(1064 纳米,0.6 W cm −2 , 10 min)时,B16F10 细胞的荧光素酶基因转染效率检测(平均值 ± SD, n = 3)。 f) pH 6.5 时,pEGFP-N1 质粒与 PAG、OEB@PAG 和 OEB@PAG+NIR-II 激光(1064 nm, 0.6 W cm −2 , 10 min)复合物处理 B16F10 细胞的流式细胞术分析。h) 不同处理后 B16F10 细胞的相对存活率(均数±标准差,n = 3)。 i) 通过流式细胞术量化 B16F10 细胞中 CRT 的暴露(均数±标准差,n = 3)。统计意义通过单因素方差分析和 Tukey 后检验计算。*p < 0.05,**p < 0.01,**p < 0.001,ns:无显著差异。
2.3 体内肿瘤靶向和生物分布
为了全面验证细菌-纳米粒子混合物EB@P在肿瘤靶向治疗中的效能与安全性,作者进行了一系列体内实验,重点关注其生物分布特性及光热响应能力。通过向B16F10肿瘤模型小鼠静脉注射Cy5.5标记的PAP复合物与EB@P混合物,并监测不同时间点的荧光信号强度,作者系统评估了它们的肿瘤靶向效率。如图4a所示,EB@P组在注射后24小时内于肿瘤区域的荧光信号显著强于PAP组,直观展示了EB@P卓越的肿瘤富集能力。
进一步分析(图4b)揭示,注射后24小时,EB@P在肿瘤组织中的荧光信号强度远超其他主要器官,这一发现不仅巩固了EB@P在肿瘤靶向方面的优势,还预示了其作为靶向药物载体的巨大潜力。此外,作者观察到EB@P在肿瘤部位的积累随时间推移而增强,并于注射后10小时达到峰值(图S17),这一时间点被确定为实施NIR-II光照射的最佳窗口。随后,红外热成像实验(图4c)在0.4 W/cm²的安全功率密度下,成功展示了EB@P在肿瘤部位的近红外-II光响应光热效应,证实其能够在体内实现温和且有效的光热加热,肿瘤部位温度达到约42.9°C(图S18)。
为了深入理解EB@P在体内的动态行为,作者对注射了EB@P的B16F10小鼠进行了时间依赖性的组织分布研究。通过收集主要器官和肿瘤组织,进行匀浆、稀释并在LB平板上培养,作者定量分析了各组织中EB@P的细菌菌落数。结果显示,在心脏、肝脏、脾脏、肺脏和肾脏等器官中,EB@P的菌落数量随时间逐渐降低,而在肿瘤组织中则呈现出先增后减的趋势,于注射后一天达到高峰(图4d,e;图S19)。这一发现表明,EB@P能够利用细菌向缺氧及免疫抑制性肿瘤微环境(TME)迁移的天然特性,在体内实现高效且持久的肿瘤蓄积。
值得注意的是,即使在注射后21天,由于免疫系统的有效清除作用,主要器官中仅检测到极少量的EB@P细菌,这充分保障了该杂交系统在临床应用中的生物安全性。综上所述,EB@P不仅展现出了优异的肿瘤靶向能力和光热响应特性,还具备高度的生物相容性和体内清除效率,为肿瘤靶向治疗提供了一种创新且高效的策略。
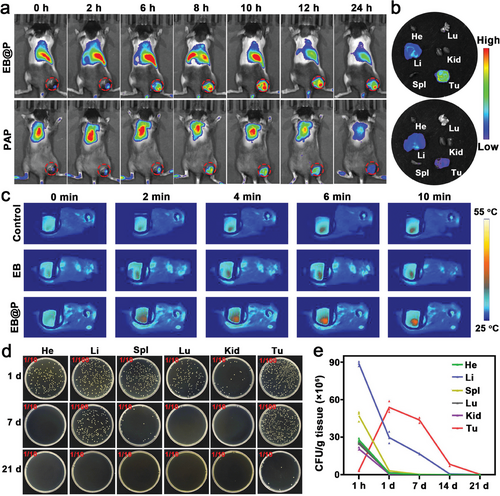
图 4 a) B16F10 肿瘤小鼠体内荧光图像;b) 静脉注射 PAP 或 EB@P 后 B16F10 肿瘤小鼠离体主要器官图像;c) 肿瘤小鼠随时间变化的光热图像。 d) 实体 LB 琼脂平板的照片;e) 在静脉注射 EB@P 后的不同时间点从肿瘤小鼠身上收集的主要器官和肿瘤中细菌定植的定量。(平均值 ± SD,n = 3)。He、Li、Spl、Lu、Kid 和 Tu 分别代表心、肝、脾、肺、肾和肿瘤。
2.4 体内抗肿瘤疗效和生物安全性评价
基于EB@P杂交种在体外展现出的强大抗肿瘤活性、低氧靶向性及良好的生物安全性,作者深入探究了其在B16F10黑色素瘤小鼠模型中的体内治疗效果。实验中,作者设计了一系列处理组,包括对照组(PBS)、仅EB+NIR-II光照射组、携带非转染细菌的OEB@PF组、EB@P+NIR-II光照射组以及集成了p53基因转染与NIR-II光响应的EB@PF+NIR-II组,以全面评估EB@P的体内抗肿瘤潜力。
如图5a所示,实验流程包括静脉注射EB@P混合物,并于注射后10小时实施近红外-II光照射。通过定期监测小鼠的肿瘤体积与体重变化(图5b,c),作者发现PBS对照组肿瘤迅速增长,符合恶性黑色素瘤的典型生长特征。相比之下,EB+NIR-II组对肿瘤生长的抑制作用微弱,表明单独的EB与NIR-II光照射对肿瘤的影响有限。OEB@PF组则显著抑制了肿瘤生长,这主要归因于p53基因工程对肿瘤细胞的直接抑制作用。
尤为引人注目的是,EB@P+NIR-II组通过结合细菌的光热响应与肿瘤免疫原性细胞死亡(ICD)机制,进一步增强了抗肿瘤效果。而EB@PF+NIR-II组则凭借p53基因转染与NIR-II光激活细菌基因工程的协同作用,实现了更为显著的肿瘤抑制效率(协同度S>0),这强调了温和光热加热对p53转染促进肿瘤细胞凋亡及PD-L1下调的关键作用(图5d,e,f)。
组织学与免疫组化分析进一步证实了EB@PF+NIR-II组的卓越疗效,表现为最广泛的肿瘤细胞坏死与凋亡,以及IL-2、p53蛋白的显著上调和PD-L1的显著下调(图5g,h,i)。这些发现不仅揭示了EB@PF杂交种在诱导抗肿瘤免疫应答方面的潜力,还强调了基因工程与光热治疗的协同增效作用。
在安全性评估方面,作者观察到各组小鼠在治疗期间体重稳定,未出现显著下降(图6a)。生化指标检测显示,肝脏(AST、ALT)与肾脏(UREA/CREA)功能均保持正常(图6b,c),表明EB@PF杂交种对全身毒性影响较小。此外,主要器官的H&E染色结果及细菌清除情况(图6d, 图S20)均证实了EB@PF在体内的高效清除与良好的生物安全性。
综上所述,EB@PF杂交种在B16F10黑色素瘤小鼠模型中展现了卓越的抗肿瘤疗效与生物安全性,为细菌介导的癌症治疗提供了有力的实验依据与潜在的临床应用前景。
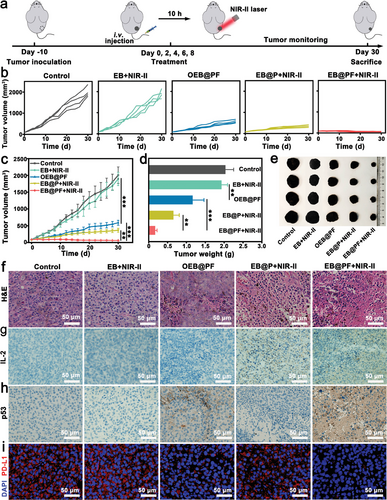
图 5 a) EB@P 介导的抗肿瘤治疗过程示意图。 b) 不同治疗后小鼠单个肿瘤的生长曲线。d) 不同治疗后小鼠平均肿瘤重量(平均值±标准差,n = 4)和 e) 不同治疗后小鼠切除肿瘤的代表性照片。 f) 不同治疗后小鼠肿瘤切片的代表性 H&E 染色图像。g) IL-2 蛋白和 h) p53 蛋白的免疫组化分析。 i) 不同治疗后肿瘤切片中 PD-L1 的免疫荧光染色。数据以平均值 ± SD 表示。统计意义采用 Tukey 后检验的单因素方差分析。**P < 0.01,**P < 0.001。
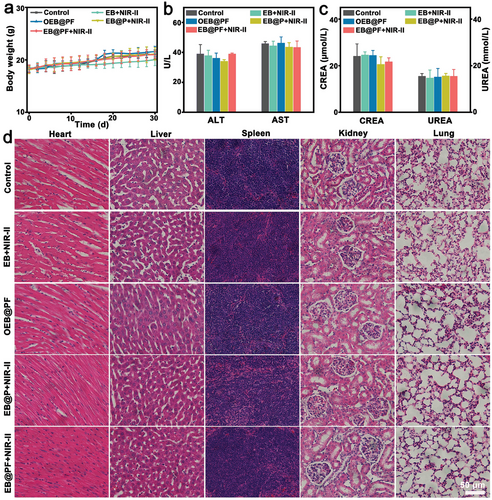
图 6 a) B16F10 肿瘤小鼠经不同处理后的体重变化(均数±标准差,n = 4);b) 与活体功能相关的酶浓度 ALT 和 AST(均数±标准差,n = 3);c) 与肾功能相关的标志物 CREA 和 UREA(均数±标准差,n = 3);d) 不同处理后小鼠主要器官(心、肝、脾、肾和肺)的 H&E 染色。
2.5 体内抗肿瘤免疫反应
为了深入探究近红外诱导的体内免疫原性细胞死亡(ICD)在激活抗肿瘤免疫反应中的关键作用,特别是其对招募与成熟抗原提呈细胞(如树突状细胞,DC)以激活T细胞的重要性,作者进行了CRT(钙网蛋白)免疫荧光染色分析,以评估不同处理组肿瘤中ICD的诱导情况。如图7a所示,经近红外-II光照射处理的肿瘤组织(EB@P+近红外-II组与EB@PF+近红外-II组)中,CRT暴露显著增加,这一发现与先前研究一致,即温和的光热加热能够有效诱导ICD,进而促进CRT的暴露。
为了进一步阐明EB@PF所介导的显著抗肿瘤效应背后的免疫学机制,作者系统地收集了淋巴结、肿瘤及脾脏中的免疫细胞,并检测了血清中的主要细胞因子。首先,利用流式细胞术对淋巴结中DC的成熟状态进行了评估。如图7b所示,与PBS对照组相比,EB+NIR-II组即使在缺乏特定基因修饰的情况下,也观察到了DC的显著成熟,这揭示了细菌本身具有的免疫佐剂特性。而OEB@PF组则表现出更高的DC成熟激活水平,这可能与基因工程赋予的免疫调节功能有关。值得注意的是,EB@P+NIR-II组与EB@PF+近红外-II组在NIR-II光照射下,DC的成熟度进一步提升,分别达到约46.4%和46.7%,这强烈提示了光热效应与基因工程协同作用下抗原释放对DC成熟的促进作用。
接下来,作者量化了肿瘤和脾脏中的T细胞亚群,包括CD4+辅助性T细胞和CD8+细胞毒性T淋巴细胞(CTLs),以评估系统性抗肿瘤免疫应答的激活情况(图7c及图S21)。结果显示,EB+NIR-II组因细菌的存在而显著增加了CTLs的浸润,但OEB@PF组尽管DC成熟度较高,CTLs浸润却未显著增加,这可能与缺乏肿瘤相关抗原(TAA)的释放有关。相反,EB@P+NIR-II组与EB@PF+近红外-II组在NIR-II光触发下,TAA的有效释放与IL-2的表达共同激活了强大的抗肿瘤免疫反应,导致CTLs浸润的显著增加。脾脏中CD4+T细胞和CTLs的活化也进一步证实了EB@PF在介导强效抗肿瘤免疫方面的潜力(图S21)。
此外,为了验证IL-2在促进自然杀伤细胞(NK细胞,CD3-NK1.1+)增殖与活化中的作用,作者分析了NK细胞在肿瘤和脾脏中的分布。如图7d及图S22所示,EB@PF+近红外-II组在这两个部位均表现出最高的NK细胞比例,表明CTLs与NK细胞均被有效激活,共同参与了抗肿瘤免疫应答。
最后,通过酶联免疫吸附试验(ELISA),作者检测了血清中多种细胞因子的水平,包括白细胞介素-6(IL-6)、γ干扰素(IFN-γ)和肿瘤坏死因子-α(TNF-α)。结果显示,EB@PF+近红外-II组这些细胞因子的水平均显著升高,进一步证实了其诱导的强烈抗肿瘤免疫反应(图7e-g)。综上所述,EB@PF混合物在促进DC成熟、CTLs浸润及NK细胞活化方面展现出卓越性能,这为其实现显著的抗肿瘤效果提供了坚实的免疫学基础。
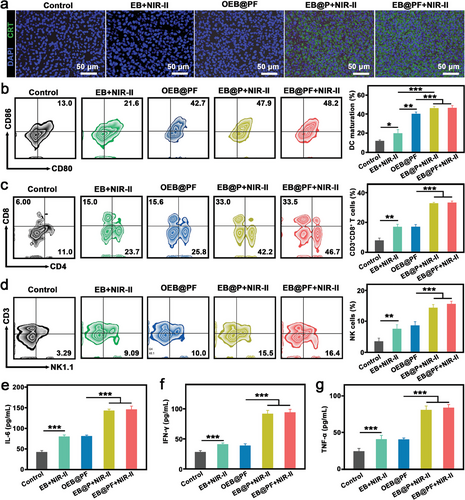 图 7
图 7
b) 淋巴结中成熟 DCs(CD11c + CD80 + CD86 + )的代表性流式细胞仪图和定量分析(平均值±标度,n = 3)。c) 肿瘤中 CD3 + CD4 + 和 CD3 + CD8 + T 细胞的典型流式细胞仪图和定量分析(平均值±标清,n = 3)。 d) 肿瘤中 CD3 − NK1.1 + NK 细胞的典型流式细胞仪图和定量分析(平均值±标清,n = 3)。通过 ELISA 分析血清中 e) IL-6、f) IFN-γ 和 g) TNF-α 的表达水平(均数±标度,n = 3)。统计意义通过单因素方差分析和 Tukey 后检验计算。*P < 0.05,**P < 0.01,**P < 0.001。
2.6 抑制远处肿瘤生长
在EB@PF诱导的显著系统性抗肿瘤免疫反应的基础上,作者进一步构建了一个远处肿瘤模型,旨在深入探索其对于抑制肿瘤复发与转移的潜在疗效(图8a)。该模型通过在小鼠右侧腹部注射B16F10肿瘤细胞作为原发肿瘤,随后在左侧腹部接种远处肿瘤来模拟临床上的肿瘤转移情境。实验结果显示,EB@PF结合NIR-II光照射处理不仅显著抑制了原发肿瘤的生长,这一趋势与先前研究(图5c)相吻合,而且EB@PF+NIR-II组展现出了最强的肿瘤抑制效果(图8b)。尤为引人注目的是,相较于EB@P+NIR-II组和OEB@PF组,EB@PF+NIR-II治疗组在抑制远处肿瘤体积增长方面表现出色(图8c),这有力地证明了其卓越的腹腔抗肿瘤效能以及细菌与肿瘤细胞基因工程联合策略的优越性。
作者推测,EB@PF在NIR-II光照射下对原发肿瘤的有效治疗,触发了强烈的抗肿瘤免疫反应,这种免疫反应进而能够清除远处的肿瘤细胞,体现了免疫治疗的远隔效应。
此外,生存分析显示,EB@PF+NIR-II组双侧黑色素瘤小鼠的生存期显著延长(图8d),且在治疗过程中小鼠未表现出明显的副作用(图S23,补充资料),进一步证实了该治疗策略的安全性与有效性。综上所述,EB@PF介导的肿瘤治疗策略在抑制远处肿瘤生长、预防肿瘤复发与转移方面展现出巨大潜力。
进一步分析免疫细胞浸润情况发现,EB@P+NIR-II组和EB@PF+NIR-II组在远处肿瘤和脾脏中的CD4+和CD8+ T细胞浸润均显著增加,这标志着抗肿瘤免疫应答的系统性激活(图8e及图S24,补充信息)。特别地,EB@PF+NIR-II组在远处肿瘤和脾脏中的CD8+ T细胞比例分别是OEB@PF组的2.2倍和2.5倍,凸显了NIR-II光触发的ICD(免疫原性细胞死亡)与细菌基因工程在激发抗肿瘤免疫中的核心作用。
同时,EB@PF+NIR-II组NK细胞在远处肿瘤和脾脏的分布也显著增多(图8f及图S25,补充资料),这验证了IL-2在促进NK细胞增殖与活化中的关键作用。此外,EB@PF介导的全身免疫反应还促进了TNF-α、IL-6和IFN-γ等细胞因子的分泌(图8g-i),这些细胞因子在抗肿瘤免疫中发挥着重要作用。
综上所述,EB@PF杂交体作为一种创新的癌症免疫疗法,通过激发强有力的抗肿瘤免疫反应,有效抑制了远处肿瘤的生长,为肿瘤复发与转移的防治提供了新的策略与希望。
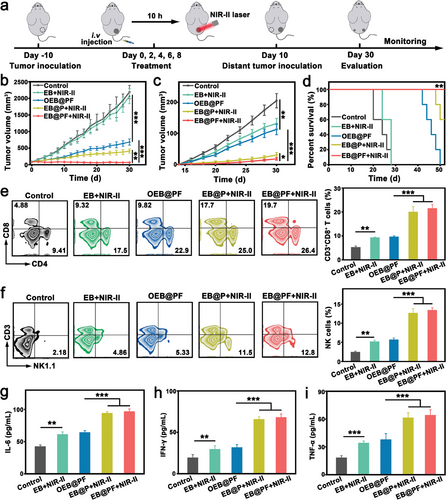
图 8 a) 治疗设计示意图。b) B16F10 肿瘤小鼠原发肿瘤 c) 和远处肿瘤经不同治疗后的肿瘤生长曲线(平均值±标度,n = 5)。 d) 肿瘤小鼠经不同治疗后的生存曲线。e) 远处肿瘤中 CD3 + CD4 + 和 CD3 + CD8 + T 细胞的典型流式细胞仪图和定量分析(平均值±标度,n = 3)。 f) 远处肿瘤中 CD3 − NK1.1 + NK 细胞的典型流式细胞仪图和定量分析(平均值±标度,n = 3)。通过 ELISA 分析血清中 g) IL-6、h) IFN-γ 和 i) TNF-α 的表达水平(均数±标度,n = 3)。统计显著性由单向方差分析和 Tukey 后检验计算得出。*ssp < 0.05,**p < 0.01,**p < 0.001。
3 结论
综上所述,综上所述,作者成功地设计并作者成功地设计并构建了基于基因工程细菌的近红外-构建了基于基因工程细菌的近红外-II光响应性EB@PF杂交体系,该体系通过巧妙地在细菌II光响应性EB@PF杂交体系,该体系通过巧妙地在细菌表面共轭光热纳米粒子得以实现。表面共轭光热纳米粒子得以实现。这一创新设计不仅利用了缺氧靶向工程菌的特性,确保了杂交种能够特异性这一创新设计不仅利用了缺氧靶向工程菌的特性,确保了杂交种能够特异性地在肿瘤部位富集,而且通过纳米粒子的光热效应,使杂交体系地在肿瘤部位富集,而且通过纳米粒子的光热效应,使杂交体系能够精准响应近红外-II光照射。能够精准响应近红外-II光照射。在光照下,温和的热效应不仅触发了免疫原性细胞死亡在光照下,温和的热效应不仅触发了免疫原性细胞死亡(ICD)的诱导,还激活了细菌内嵌的基因工程机制。(ICD)的诱导,还激活了细菌内嵌的基因工程机制。
尤为值得一提的是,通过NIR-II光的激活,作者实现了目标治疗性免疫尤为值得一提的是,通过NIR-II光的激活,作者实现了目标治疗性免疫调节因子IL-2的表达,这一过程极大地促进了细胞毒性T淋巴细胞调节因子IL-2的表达,这一过程极大地促进了细胞毒性T淋巴细胞(CTLs)的增殖与活化,为抗肿瘤免疫反应注入了强大动力。(CTLs)的增殖与活化,为抗肿瘤免疫反应注入了强大动力。此外,作者还巧妙地利用了希夫键合的pH响应性,在肿瘤此外,作者还巧妙地利用了希夫键合的pH响应性,在肿瘤微环境(TME)的特定酸性条件下,释放出装载有肿瘤细胞基因微环境(TME)的特定酸性条件下,释放出装载有肿瘤细胞基因工程所需纳米颗粒的载体,实现了对肿瘤细胞的精准基因调控。工程所需纳米颗粒的载体,实现了对肿瘤细胞的精准基因调控。
特别地,抗原p53在B16F10肿瘤细胞中的特别地,抗原p53在B16F10肿瘤细胞中的表达策略不仅有效诱导了肿瘤细胞的凋亡,还通过下调程序性死亡表达策略不仅有效诱导了肿瘤细胞的凋亡,还通过下调程序性死亡配体1(PD-L1)的表达,显著削弱了TME中的配体1(PD-L1)的表达,显著削弱了TME中的免疫抑制效应,为免疫细胞的浸润与功能发挥创造了有利条件。免疫抑制效应,为免疫细胞的浸润与功能发挥创造了有利条件。
更为显著的是,作者的研究揭示了细菌基因工程与肿瘤细胞基因工程之间的协同作用更为显著的是,作者的研究揭示了细菌基因工程与肿瘤细胞基因工程之间的协同作用,这种双重基因工程策略在抗肿瘤治疗中展现出了强大的级联增强,这种双重基因工程策略在抗肿瘤治疗中展现出了强大的级联增强效应,能够显著抑制原发性和远处肿瘤的生长。效应,能够显著抑制原发性和远处肿瘤的生长。同时,EB@PF杂交种所表现出的优异生物安全性,为其在未来体内同时,EB@PF杂交种所表现出的优异生物安全性,为其在未来体内肿瘤治疗中的广泛应用奠定了坚实基础。肿瘤治疗中的广泛应用奠定了坚实基础。
综上所述,本项工作不仅为细菌-纳米粒子杂交平台的合理设计与构建综上所述,本项工作不仅为细菌-纳米粒子杂交平台的合理设计与构建提供了新思路,更为开发安全高效的癌症免疫治疗策略开辟了新途径,提供了新思路,更为开发安全高效的癌症免疫治疗策略开辟了新途径,具有重要的理论意义与广阔的应用前景。具有重要的理论意义与广阔的应用前景。